
QUBIC BLOG POST
What Can a Neuron Compute? Dendritic Computation and Brain-Inspired AI
Written by

Qubic Scientific Team
Published:

Listen to this blog post
An analysis by the Qubic Scientific Team of "What can a neuron compute" (Aizenbud et al., 2026)
What a Biological Neuron Knows How to Do, and Compute
A neuron receives signals, integrates them and, if the integration exceeds a threshold, emits a signal of its own. The dendrites are the branches that receive the input. The soma is the body where, in the simplified version, everything is summed. The axon is the output line that transmits the spike. The contact points are the synapses, and their strength changes with learning.
For half a century, artificial intelligence worked with this scheme. The McCulloch–Pitts neuron, and later Rosenblatt's perceptron (Rosenblatt, 1958), reduce everything to a weighted sum followed by a threshold. It is a convenient and, in a sense, fertile abstraction: by stacking millions of these units one builds the networks that today dominate AI. But that point-neuron cannot solve, on its own, problems that are not linearly separable. XOR, the canonical example, lies beyond its reach. The problem is that the biological neuron was never like that. It receives thousands of synapses distributed across a branched dendritic tree, and those inputs interact nonlinearly at many different places before reaching the soma. For years the contribution of the individual cell to the brain's computation was underestimated, precisely because it was taken for granted that all the richness lived in the network and not in its units (London & Häusser, 2005).
The Cortex and Its Layers: Where Cortical Computation Lives
The neuron that interests us lives in a specific place. The neocortex, where much of cognitive computation takes place, is organized into layers: conventionally six are counted, stacked from the surface inward. Each layer has its own profile of inputs and outputs; the upper ones receive and distribute information within the cortex, the deep ones send it outward. Layer 5 occupies a privileged place in this scheme. It is, to a large extent, the output layer, and it hosts some of the largest and morphologically most complex cells in the entire cortex. That these neurons extend their dendrites across several laminae is no accident: it lets them couple information arriving at different cortical heights, a cellular mechanism described a quarter of a century ago now (Larkum, Zhu & Sakmann, 1999). If we want to know how much a single neuron can compute, this is a good place to look.
The Layer 5 Pyramidal Neuron (L5PC): A Two-Layer Network in a Single Cell
The protagonist of the paper we are discussing is the layer 5 pyramidal cell, the L5PC. Its shape gives it its name. From the soma, with its roughly triangular body, two dendritic systems emerge. The basal dendrites extend around the soma. And a large apical dendrite ascends toward the cortical surface and opens into a tuft that reaches layer 1. This physical separation is not trivial: apical and basal receive inputs from different sources and can operate as semi-independent subunits.
What is decisive is that these dendrites are not passive. They contain active conductances that generate local nonlinearities (NMDA spikes; Schiller, Major, Koester & Schiller, 2000). Each branch can, in effect, perform a small nonlinear operation before its result reaches the soma. Hence the classic proposal of reading the pyramidal cell as a two-layer network: the branches compute local functions and the soma integrates them (Poirazi, Brannon & Mel, 2003).
What the TwinProp Paper Does, and Why Single-Neuron Computation Matters
Optimizing at once the strength and the position of thousands of synapses on a detailed biophysical model is costly. Without a tool to search for good synaptic configurations, there was no way to know whether the failures at solving problems like XOR were limits of the neuron or limits of the method.
TwinProp solves this with an elegant detour (Aizenbud et al., 2026). Since the detailed model is not conveniently differentiable, the authors train a differentiable digital twin that mimics its behavior at the millisecond, taking advantage of the fact that the input–output relationship of a layer 5 pyramidal cell can be captured with a deep neural network (Beniaguev, Segev & London, 2021). On that twin they can indeed backpropagate gradients and find the synaptic weights that solve a task; they then transfer those weights to the detailed neuron and check that they work. It is worth stressing a nuance the paper itself is careful about: TwinProp is an instrument for discovering what the neuron can compute, not a model of how it learns biologically.
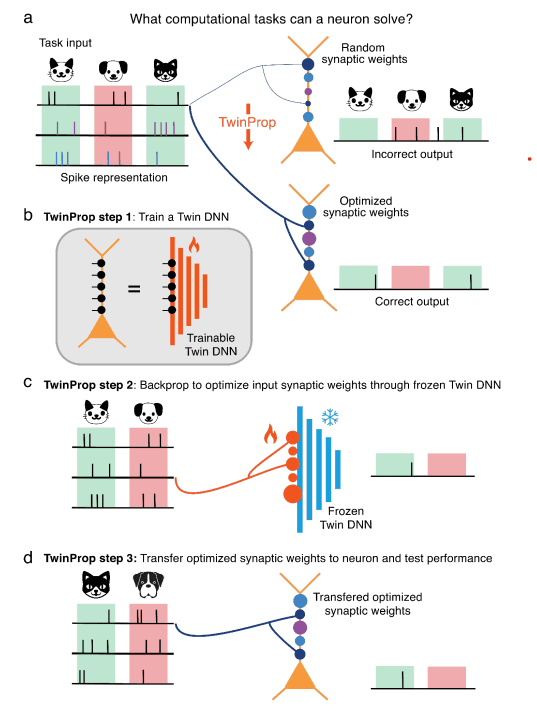
Figure 1. The TwinProp algorithm for probing the range of tasks a single neuron can implement. Reproduced from Aizenbud et al. (2026), licensed under CC-BY-NC-ND 4.0.
The results are compelling. A single L5PC classifies images and voice commands above the linear models, and solves arbitrary Boolean functions with accuracy close to the ceiling of a deep decoder. These are problems the textbook attributes to multilayer networks, solved by one cell. And the why is mechanistic: the greater the difficulty, the more the distributed dendritic nonlinearities are recruited, especially the NMDA-mediated ones, and the higher the rank of the voltage dynamics in the tree. Complexity is an ornament.
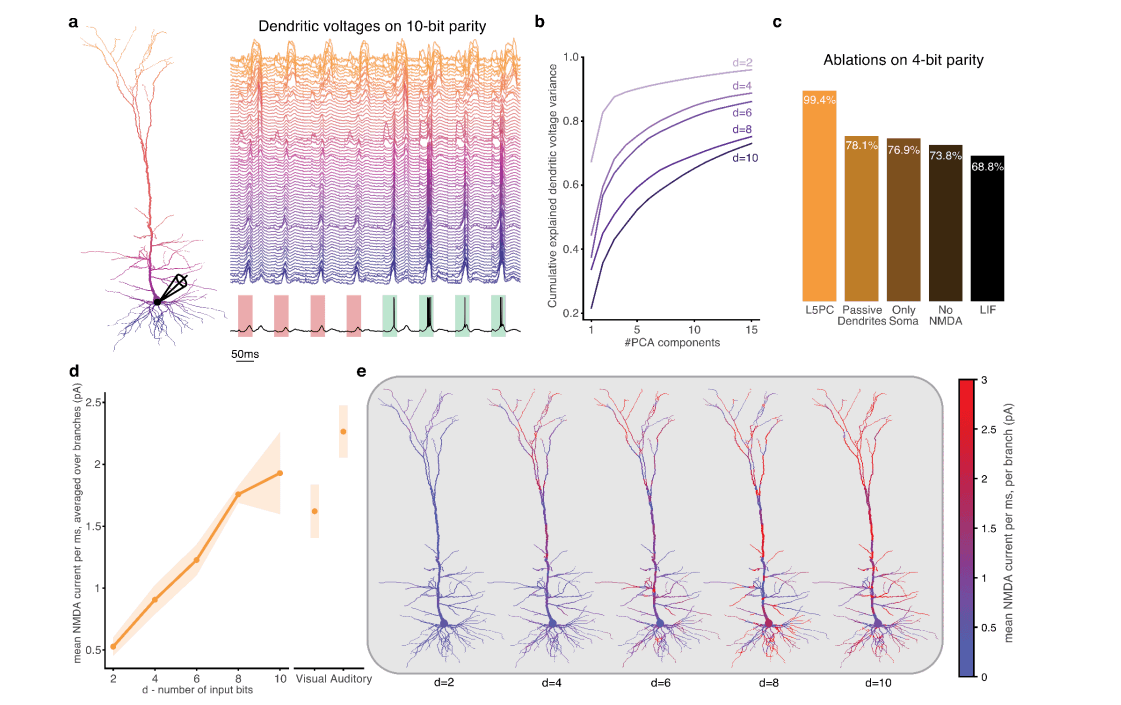
Figure 2. Distributed, NMDA-amplified dendritic integration underlies single-neuron solutions to hard nonlinear tasks. Reproduced from Aizenbud et al. (2026), licensed under CC-BY-NC-ND 4.0.
The relevance goes beyond cellular neuroscience. Modern artificial intelligence was built on the bet that the individual neuron could be abstracted away without loss, letting power emerge from number and quantity. This work quantifies the cost of that bet. If a single cell binds high-order features thanks to its structure, replacing it with a sum plus a threshold discards real computation. The ability to mix many task variables nonlinearly is, moreover, precisely the property that gives high dimensionality to neural representations and broadens the repertoire of functions that a downstream reader can implement (Rigotti et al., 2013). It is not an argument against deep networks, but about what is lost by choosing the poorest possible unit as the building block.
Neuraxon: Time, Neuromodulation, and Bio-Inspiration
This is where the paper enters into conversation with Neuraxon.
Bio-Inspiration: Computation Lives in the Structure of the Cell
The underlying parallel is bio-inspiration. The paper's thesis is that computational richness lives in the structure of the cell, not only in the network. It is exactly the premise we start from. We do not treat the Neuraxon as a point that sums and fires; we treat it as a unit whose internal structure performs computation. That a detailed biophysical model outperforms the perceptron by a wide margin reinforces, from neuroscience, the design decision we made from engineering. (For a broader comparison of biological and artificial neural networks, see NIA Volume 4: Neural Networks in AI and Neuroscience.)
Time as a First-Class Variable in Neural Computation
The second parallel is time. The neuron in the paper does not classify by mean firing rates: it exploits the fine temporal structure of the spikes, and its solution degrades gracefully when temporal noise is added, which reveals that the exact timing of each spike carries information. That idea has firm roots: it had already been shown that cortical neurons generate reproducible spike trains with sub-millisecond precision when they receive realistic fluctuating inputs (Mainen & Sejnowski, 1995). In our case time is not a side effect but a first-class variable. Continuous-time processing and time-embedded state dynamics make the when part of the computation, not an implementation detail. The difference is one of degree and of intention: they discover that their neuron uses time; we build it as a principle. (We develop this fully in NIA Volume 1: Why Intelligence Is Not Computed in Steps, but in Time.)
Neuromodulation: From a Static Neuron to a Reconfigurable One
The third piece marks a clear distance: neuromodulation. In the paper, once optimized, the synaptic weights remain fixed. The neuron is powerful, but static. Neuromodulation, which in the brain reconfigures the gain and plasticity of entire circuits according to the organism's state, is not part of the model. We incorporate it as an active mechanism: the Neuraxon's computation can be reconfigured on the fly.
The paper demonstrates computational capability in a simulated cell; nothing in it touches consciousness. We claim it nor disguise it with pseudo-quantum jargon. Neuraxon is a computational project. If power lives in structure, in time and, in biology, in the modulation of state, then an architecture that starts from those three principles is not just one more alternative among many. Another small angle for understanding that Qubic's ideas, far from being random, are well rooted in biology.
Scientific References
Aizenbud, I., Beniaguev, D., Pnueli, N., Segev, I., & London, M. (2026). What can a neuron compute. bioRxiv. https://doi.org/10.64898/2026.06.08.730984
Beniaguev, D., Segev, I., & London, M. (2021). Single cortical neurons as deep artificial neural networks. Neuron, 109(17), 2727–2739.e3.
Larkum, M. E., Zhu, J. J., & Sakmann, B. (1999). A new cellular mechanism for coupling inputs arriving at different cortical layers. Nature, 398(6725), 338–341.
London, M., & Häusser, M. (2005). Dendritic computation. Annual Review of Neuroscience, 28, 503–532.
Mainen, Z. F., & Sejnowski, T. J. (1995). Reliability of spike timing in neocortical neurons. Science, 268(5216), 1503–1506.
Poirazi, P., Brannon, T., & Mel, B. W. (2003). Pyramidal neuron as two-layer neural network. Neuron, 37(6), 989–999.
Rigotti, M., Barak, O., Warden, M. R., Wang, X.-J., Daw, N. D., Miller, E. K., & Fusi, S. (2013). The importance of mixed selectivity in complex cognitive tasks. Nature, 497(7451), 585–590.
Rosenblatt, F. (1958). The perceptron: A probabilistic model for information storage and organization in the brain. Psychological Review, 65(6), 386–408.
Schiller, J., Major, G., Koester, H. J., & Schiller, Y. (2000). NMDA spikes in basal dendrites of cortical pyramidal neurons. Nature, 404(6775), 285–289.